18
Faunal Studies
Jonathan C. Driver, Michael J. Brand, Lianne Lester, and Natalie D. Munro
Contents
Identification and Data Recording
- Taxa Identified
- Amphibians and Reptilesbr /> Birdsbr /> Mammals
- Lagomorphsbr /> Rodentsbr /> Carnivoresbr /> Artiodactylsbr /> Perissodactyl
Explanation of Intersite Variation
Introduction
This chapter provides a broad overview of zooarchaeological studies undertaken on 13 sites excavated as part of the Sand Canyon locality Site Testing Program. The objectives of this chapter are (1) to assess the major factors which have contributed bones to the recovered assemblages and (2) to examine variation in assemblage composition between sites and attempt to explain such variation. Other faunal studies in the Sand Canyon locality include recently completed analyses of turkey bone (Munro 1994) and bone tools (Bullock 1992). Walker analyzed faunal remains from the early seasons of excavation at Sand Canyon Pueblo (Walker 1990a) and from the Green Lizard site in Sand Canyon (Walker 1990b); he also reported the fauna from the Pueblo I Duckfoot site, just outside the Sand Canyon locality (Walker 1989, 1993). Brand (1991) examined other faunal assemblages from Sand Canyon Pueblo. Ongoing faunal work includes a reexamination and spatial analysis of the entire Sand Canyon Pueblo faunal assemblage and a comparative study of faunal remains from the Mesa Verde region. This chapter begins with a brief description of the methods used in recording the faunal assemblages, followed by a discussion of the taxa identified. Unlike other chapters in this volume, we do not discuss individual site assemblages in detail, because the taxa identified are similar from site to site and because variation is most easily seen between groups of sites. Furthermore, many assemblages are too small to provide any useful data on intrasite variability and can only be interpreted by grouping certain site assemblages together and then comparing these groups to one another. Site descriptions and locations can be found in Chapters 2 through 14 of this volume.
Identification and Data Recording
All nonhuman bones recovered from the tested sites are discussed in this chapter. These include two classes of faunal remains: modified nonhuman bone (that is, bone artifacts and the by-products of artifact manufacture) and unmodified nonhuman bone (that is, animal bones which were not deliberately shaped by humans, but which might have been modified in other ways by humans, such as by burning or butchering, or by natural processes such as weathering or rodent gnawing). This chapter combines data from both classes of bone, such that the total number of bones includes worked and unworked specimens. Bone artifacts are discussed in more detail in Chapter 15 of this volume.
Standard information was recorded for each specimen, using coding procedures devised by Driver and described in a manual on file at the Crow Canyon Archaeological Center. The following information was recorded for each specimen. Provenience was recorded following the Crow Canyon numbering system. Taxon was determined for most specimens using the Simon Fraser University zooarchaeological collection, with the assistance of published guides, notably those of Olsen (1964) and Lawrence (1951). A few specimens were identified at the University of Puget Sound Museum of Zoology. Taxon identifications ranged from the species level to "unidentified." Element was defined using standard zoological terminology. For all specimens in which the element could not be identified, the taxon was automatically defined as "unidentified," following a rationale discussed by Driver (1992). Part of element was coded to allow some detail in the description of which part of a bone was present. This ranged from very simple descriptions for some bones (for example, sesamoids were recorded as either complete or fragmentary) to more complex descriptions for others (for example, long bones could be assigned one of 11 different codes depending on which part of the bone was present). Side of the body was recorded. When appropriate, the state of fusion of epiphyses was recorded to assist in determination of the age structure of the various animal populations represented in the sites. Breakage (the way in which bones were broken at either end) was assessed for every specimen, using description of fracture type. Any modification to the bone was noted. Modifications included animal modification (for example, carnivore or rodent damage) and human modification (for example, cut marks, modification as an artifact, burning). The length of every fragment was recorded to the nearest centimeter. The cortical bone thickness of all fragments with a tubular structure (that is, long bone fragments) was also recorded to the nearest 2-mm increment. All bone which was deliberately modified by humans was separated for further study. Any human bone identified during analysis was also separated and returned to Crow Canyon. All faunal data are stored in a master database at Crow Canyon.
The wide variety of Sciuridae (chipmunks, ground squirrels, prairie dogs, etc.) are difficult to separate osteologically. Most identifications to genus and species have been made on mandibles and crania. Many specimens of long bones and axial fragments have been identified only to the family level. No attempt has been made in this study to distinguish domestic from wild turkey (although it appears that most of the specimens are domestic), nor is the identification of different breeds of domestic turkey (McKusick 1986) discussed. Munro (1994) should be consulted for more detail. A few other bird specimens eventually may be identified to more specific levels than are reported in this chapter, although the overall number of nonturkey birds is very small, and more precise identifications will not affect the general conclusions about these assemblages.
Taxa Identified
Table 18.1 summarizes all bone recovered from all 13 sites included in this study. When possible, taxa are identified by common names in Table 18.1. The following sections provide more detailed taxonomic information. As can be seen, certain numerically important taxa are consistently found in most sites, although the proportions vary, as discussed later. These common taxa typically include turkey, cottontail rabbit, jackrabbit, sciurids, pocket gopher, packrat, and artiodactyls, all of which are found in more than half of the assemblages. Rather than discuss the assemblages from each site repetitively, it is more convenient to discuss each taxon once. The range of taxa represented is consistent with the modern fauna of the area and with other faunas reported from archaeological sites in the Sand Canyon locality or nearby areas (Neusius 1986; Walker 1990a, 1990b, 1993).
Amphibians and Reptiles
Amphibians and reptiles are represented by frogs, toads, lizards, and snakes. No species identifications were made. Bones were generally found as rare isolated elements, but in two cases partly articulated skeletons were recovered--the snake from Catherine's Site and the large toad from Kenzie Dawn Hamlet. Although amphibians and reptiles may have been important in ritual, and representations of snakes are found in rock art in the region, the recovered bones probably represent animals which had entered the sites after they were abandoned by the Puebloans. There is no evidence that amphibians and reptiles were economically important in any of these sites, nor in any other area of the Southwest.
Birds
As noted by Walker (1990a, 1990b), the avian fauna from Sand Canyon locality sites is not very diverse when compared with other Southwestern assemblages. The sites reported here support Walker's conclusions. Turkey (Meleagris gallopavo) is by far the most common identified bird species. Turkey was identified on the basis of cranial elements, pectoral and pelvic girdles, and major limb bones. Because of a poor comparative collection, vertebrae, ribs, and phalanges were not often identified to species and were instead assigned to the "large bird" category. However, it is reasonable to assume that virtually all of the "large bird" bones are also from turkey, and all discussion of turkey in the remainder of this paper combines the two categories. The economic and ritual importance of turkey in Southwestern cultures is well known (Gnabasik 1981; McKusick 1986).
Other birds found at these sites include an array of Falconiformes. Large hawks (Buteo sp.) were found in four sites (Castle Rock Pueblo, Shorlene's Site, Catherine's Site, and Kenzie Dawn Hamlet), and a large falcon (Falco sp.) was also recovered from Castle Rock Pueblo. The latter specimen is morphologically very similar to peregrine falcon and only slightly smaller than the modern specimen with which it was compared. The sparrowhawk (Falco sparverius, also known as the American kestrel) was found at two sites, Kenzie Dawn Hamlet and Stanton's Site. Birds of prey had considerable significance in Southwestern ritual (Gnabasik 1981; Tyler 1991), although the eagles occupy more space in the literature than the hawks. Interestingly, no eagle bones were found in the sites discussed here, even though Falconiformes were present in about half the assemblages. The feathers of the sparrowhawk, the smallest of the Falconiformes, are specifically named in descriptions of some ceremonies (Gnabasik 1981) and may be associated with the initiation of young men (Tyler 1991).
Apart from the turkey, other Galliformes such as grouse (Tetraonidae) and quail (Phasianidae) may have been obtained for either food or feathers. The same could be said for the unidentified duck bone. The small pigeon or dove is probably the mourning dove (Zenaidura macroura), which is commonly seen in the region today. A single bone of the common flicker (Colaptes cafer) might represent an animal hunted for its feathers, as might the presence of raven (Corvus corax) and magpie (Pica pica).
Mammals
Lagomorphs
The mammalian assemblage is dominated by Lagomorpha and Rodentia. Lagomorphs include cottontails (Sylvilagus sp.) and jackrabbits (Lepus sp.). No attempt was made to identify the species represented, although Neusius and Flint (1985) have shown that it may be possible to separate Sylvilagus audubonii from Sylvilagus nutallii using measurements of the mandible. Brand (1991), using Neusius and Flint's method, demonstrated that both species were probably present in Sand Canyon locality assemblages. Although the Lepus specimens could not be assigned to a species, their size clearly excludes the possibility that Lepus americanus (snowshoe hare) was present, and they therefore represent one of two species of jackrabbit, probably Lepus californicus.
Rodents
Numerous species of rodent are present in the region today, and this is reflected in the diversity of rodents recovered from all of the sites. The squirrel family (Sciuridae) is represented by many species. Taxa identified include Eutamias sp. (chipmunk), Spermophilus sp. (ground squirrel), Spermophilus variegatus (rock squirrel), and Cynomys sp. (prairie dog). Most of the ground-squirrel bones are similar in size to rock squirrels, but two smaller mandibles from Castle Rock Pueblo and Stanton's Site are very similar to spotted ground squirrel (Spermophilus spilosoma). Most of the Sciuridae construct burrows for nests and storage, and it is therefore quite possible that they are intrusive to the archaeological deposits. This is probably the case for the chipmunk at Lester's Site. However, some are definitely edible and provide as much meat as a cottontail, and ridding fields and storage areas of these animals would enhance agricultural productivity.
Pocket gophers (Geomyidae) are difficult to identify to genus and species. The few specimens which could be identified further (based on incisor morphology) were all Thomomys. Pocket gophers burrow extensively, rendering them likely candidates for intrusive burrowing. However, they would also be serious agricultural pests and might be trapped for this reason. They would provide little meat.
Of the Heteromyidae, only Ord's kangaroo rat (Dipodomys ordii) has been identified so far. This species is likely to be intrusive.
Identified mice and voles (Cricetidae) include Peromyscus sp. and Microtus sp. Some specimens are almost certainly intrusive. For example, during excavation of Sand Canyon Pueblo, the dessicated carcass of a complete mouse was recovered, suggesting death within the last few years. Many of the small rodent bones may reflect similar natural deaths between site abandonment and excavation. Even if some small rodents were consumed, their contribution to the human diet would have been insignificant.
Packrats or woodrats (Neotoma sp.) are also attracted to archaeological sites as nesting areas, and any sites built near rocky outcrops may overlap packrat nesting sites. Packrats might be trapped for food (especially as they would have been a potential consumer of stored plant foods), and were sometimes caught during communal rabbit drives (Gnabasik 1981), but they might also be postabandonment inhabitants of the sites.
There are various ways to assess whether rodents were used as food. Although accelerator mass spectrometry (AMS) radiocarbon dating of individual bones can be used to assess the rough contemporaneity of burrowing rodents with the strata in which they are found (e.g., Driver 1988), this will not necessarily provide useful information in the later prehistoric periods of the Southwest. For example, AMS dating of an articulated rodent skeleton from a burrow in an archaeological site in New Mexico provided a date indistinguishable from dates on culturally deposited animal bone and charcoal from the same room, demonstrating that rodents may move into sites immediately after abandonment (Driver 1991). Small mammals are not expected to display much evidence of butchering (although see Patou [1987] for examples), so the presence or absence of cut marks is not significant. Localized patterned burning of bones may reveal human cooking methods (e.g., Vigne and Marinval-Vigne 1983), but this form of burning is rare in Sand Canyon locality sites, probably because meat was only rarely roasted over an open fire.
Shaffer (1992a) has suggested that pocket gophers might have been trapped by humans in the prehistoric Southwest, and proposes that high numbers of crania and mandibles and relatively low numbers of the larger postcranial bones (femur, innominate, scapula) would characterize a humanly influenced assemblage. The largest pocket gopher assemblage in the sites discussed here is from Kenzie Dawn Hamlet. Mandibles and crania account for 46 percent of the assemblage. Shaffer's data from Nan Ruin show 79 percent of the assemblage was composed of these elements. The Kenzie Dawn Hamlet assemblage does not have the extreme disparity of elements which led Shaffer to suggest that the Nan Ruin pocket gophers were collected by humans, and we conclude that the pocket gophers at Kenzie Dawn Hamlet are probably intrusive.
This conclusion is reinforced in another study by Shaffer (1992b) in which skeletons of various rodent species were screened through quarter-inch screens. He found for pocket gopher that the cranium, mandible, scapula, innominate, and femur were recovered more than 70 percent of the time, and that the sacrum would be recovered frequently because it fuses to the innominate. All other elements were recovered less than 70 percent of the time. The Kenzie Dawn Hamlet assemblage contains all of the elements expected from this study: cranium (N = 4), mandible (N = 8), scapula (N = 2), innominate (N = 5), sacrum (N = 2), and femur (N = 2). It also contains the ulna (N = 2) and the tibia (N = 2). This suggests an assemblage biased by archaeological collecting methods, but not necessarily by the effects of prehistoric trapping and consumption.
Bone breakage may provide some distinctions between intrusive and ancient animals, because intrusive animals are less likely to be affected by a wide range of taphonomic factors than bones deposited while a site was in active use. We hypothesize that intrusive specimens are more likely to be preserved as whole specimens, whereas humanly collected specimens are more likely to be broken during processing or as a result of factors such as trampling prior to burial. In this study, we compare the breakage of sciurids, packrats, and cottontails from three of the large assemblages (Table 18.2). In so doing, we are making the assumption that a significant proportion of the cottontail was consumed as food, and we are using cottontail as a standard for what consumed small mammal assemblages should look like. Bones from the three largest assemblages--Castle Rock Pueblo, Kenzie Dawn Hamlet, and Stanton's Site--have been selected for analysis.
The upper figures in Table 18.2 describe the percentage of unbroken bones identified to a particular taxon. In all sites, packrat bones are more frequently found as complete specimens than are cottontail, and in two of the three sites, sciurids are represented by relatively high percentages of complete elements. The second set of figures defines what percentage of fragment ends were coded as intact, as opposed to being broken in some way. Again, the data suggest less breakage and fragmentation of packrat and sciurid than cottontail. The figures suggest that rodents are more likely to be intrusive than are cottontails, but they do not really provide a basis for estimating the relative proportions of intrusive specimens for any group.
Another way to determine intrusiveness of rodents is to look at indices of preservation. Because many destructive taphonomic factors are cumulative (for example, weathering, trampling), one would expect intrusive specimens to suffer less than ancient specimens. One way to measure relative destruction of bone is to compare proximal and distal ends of long bones in which the ends exhibit markedly different survival properties. Here the proximal ends (fragile) and distal ends (robust) of humeri and tibiae are used. There are insufficient packrat humeri and tibiae preserved to undertake the analysis, but sciurids and cottontail rabbits can be compared (Table 18.3). Proximal ends are better represented in sciurid samples than in cottontail samples, suggesting more frequent destruction of cottontail bones. We infer that lower rates of bone destruction for rodents mean that more of them are intrusive.
Carnivores
Carnivores are found infrequently in the Sand Canyon locality sites. A spotted skunk (Spilogale putorius) was recovered from Troy's Tower, and a badger (Taxidea taxus) was recovered from Lookout House. Some of the "medium-sized carnivores" are probably mustelids. Canidae are represented by domestic dog, scattered remains identified as Canis sp. (which could be dog or coyote but are too small to be wolf), and some fox bones, of which one, from Kenzie Dawn Hamlet, could be identified positively as red fox (Vulpes vulpes). Felidae from three sites are represented by Lynx sp., which is almost certainly bobcat (Lynx rufus) on the basis of size.
Artiodactyls
Artiodactyls are rare but present at most of the tested sites. The only large artiodactyl bones, recovered from near the surface of Lillian's Site, could well be from a domestic cow. All the medium-sized artiodactyls which could be identified to genus were deer (Odocoileus). Deer is the most common artiodactyl from Sand Canyon Pueblo, although pronghorn antelope and bighorn sheep have also been recovered from that site (Brand 1991; Walker 1990a). It is possible that low percentages of artiodactyls could result from intensive bone smashing during processing, which would render more bones unidentifiable. However, an analysis of unidentified fragments does not support this hypothesis. During analysis, the cortical thickness of all long bone fragments was measured. Only 6.5 percent of all fragments have a cortical thickness greater than 2 mm, and only 1.1 percent have a thickness greater than 4 mm. As virtually all mature deer long bones have cortical thicknesses greater than 2 mm and many have a thickness greater than 4 mm, it is clear that the lack of deer bones cannot be accounted for by the "unidentified" fragments.
Element counts for all deer and medium-sized artiodactyl from the tested sites are presented in Table 18.4. The sample is too small for a taphonomic analysis. However, the number of metapodial fragments is noteworthy, especially because so many have been modified as artifacts. The relatively low percentage of artiodactyls in all assemblages and the common use of their bones as artifacts support the idea that they were infrequently procured. For example, an analysis of artiodactyls from sites in eastern New Mexico dating to the thirteenth and fourteenth centuries A.D. showed that between 2 and 16 percent of medium artiodactyl long bones were modified as artifacts (Driver 1985:58), whereas about one-third of the long bones in Sand Canyon locality sites have been modified.
Perissodactyl
Two horse teeth were identified from Lester's Site. These are interpreted as historic-period additions.
Sample Size and Taphonomy
Sample Size
Before attempting comparisons which might produce data relevant to understanding prehistoric cultures, it is necessary to consider sample size and preservation. Sample size affects the diversity of assemblages, such that smaller assemblages generally contain fewer taxa than larger assemblages. Biologists have recognized this phenomenon for many years (Fisher et al. 1943). Relatively few archaeologists seem aware of the problem, although it has been discussed at length by Leonard (1989) for prehistoric Puebloan faunas. Assemblage diversity for Sand Canyon locality assemblages was calculated by counting the minimum number of taxa required to account for the identified bones. Thus, "large bird" would not be included in the taxa counts if turkey was identified, because all large bird bones could be from turkey. Similarly "rodent" would not be counted as a separate taxon if other rodent taxa (for example, packrat, vole) had been identified. Plotting sample size against number of identified taxa (Figure 18.1) demonstrates that the expected relationship holds for the Sand Canyon locality assemblages. This means that caution must be used when comparing assemblages. The assemblage from Mad Dog Tower is so small that comparison with other sites cannot be justified. Three sites--Castle Rock Pueblo, Kenzie Dawn Hamlet, and Stanton's Site--produced the three largest assemblages (885 to 1,058 identified specimens); the remaining nine sites produced between 72 and 398 identified specimens. Two of the large assemblages--Castle Rock Pueblo and Kenzie Dawn Hamlet--exhibit the expected diversity. The assemblage from Stanton's Site has a rather low diversity for its size. This may be because Stanton's Site is a single-component, single-household site, whereas the other two sites which produced large assemblages are either multicomponent sites or housed larger populations (see Chapter 5, Kenzie Dawn Hamlet; Chapter 10, Stanton's Site; and Chapter 14, Castle Rock Pueblo).
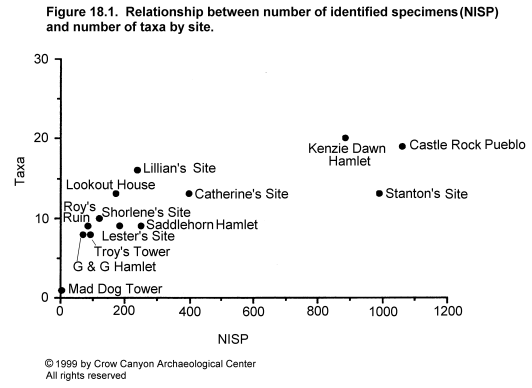{kind=link}
One must also bear in mind that sample sizes of identified bones for many sites are very small. As a result, detailed analyses of even the common taxa are difficult.
Taphonomy
We know a good deal about the effects of many natural and cultural processes on survival, modification, and destruction of bone (Lyman 1994). However, we have been less successful at understanding the cumulative effect of different processes acting on the same assemblage, either simultaneously or sequentially.
In this analysis, our aim is to assess in a relative way the extent to which penedepositional and postdepositional conditions (perthotaxic and taphic factors) may have varied from one site to another. We assume that human treatment of bones did not differ drastically from one site to another and that differences in preservation will result from "natural" conditions, such as depth of deposits, weathering rates, water percolation, etc. To compare preservation between assemblages, we have developed a series of indices summarized in Table 18.5. These are discussed in turn below.
The percentage of unidentifiable bones in an assemblage should provide a measure of bone fragmentation, assuming that more heavily fragmented specimens are likely to be less "identifiable" than relatively complete specimens. It should be remembered that in this analysis any bone fragment which could not be assigned to an element was considered as unidentifiable. Unidentified bones make up 40 percent to 92 percent of assemblages, but the majority of assemblages contain between 40 percent and 60 percent unidentified specimens. There is a good correlation between numbers of identified and unidentified specimens, which suggests that the majority of assemblages have experienced similar breakage rates.
Average length of fragments might also reflect rates of bone fragmentation, but fragment length will also be affected by the size of the animals which dominate the assemblages. It can be seen from Table 18.5 that, at most sites, average fragment length was between 2.7 and 3.3 cm, although two sites have notably smaller average lengths.
Burnt bone survives some forms of weathering and adverse soil conditions better than unburnt bone, and high percentages of burnt bone can be correlated with other indicators of poor preservation conditions (Driver 1993). Although cultural activity (for example, burning a midden) could result in significant quantities of burnt bone in an assemblage, we attempted to assess preservation using percentage of burnt bone as a possible indicator. Our assumption is that where soil chemistry and/or groundwater destroy bone, assemblages will contain higher percentages of burnt bone. Most assemblages have between about 10 percent and 20 percent burnt bone. Three assemblages (Mad Dog Tower, Lillian's Site, and G and G Hamlet) have higher percentages of burnt bone, and it is interesting that two of these sites also had small average fragment lengths, and that one (Mad Dog Tower) had the highest percentage of unidentified bone.
Another index developed to assess preservation is based on the well-documented phenomenon that proximal and distal ends of the same bone element may have drastically different survival chances. For example, Binford (1981) plotted frequency of proximal humeri against frequency of distal humeri to determine whether assemblages had been "ravaged" by carnivores. Similarly, Brain (1981) used differential preservation of proximal and distal long bones to establish the effects of carnivores on bone assemblages. Analysis of small mammals has also used this sort of index to establish the effects of predators on assemblages (e.g., Andrews 1990; Kusmer 1990). Sylvilagus is the most common mammal at most sites, and we have calculated the ratio of proximal humerus plus proximal tibia to distal humerus plus distal tibia. In well-preserved assemblages, the ratios should be approximately equal. In poorly preserved assemblages, there should be more distal than proximal ends. For many sites, the sample was too small to calculate the ratio. Table 18.5 reports data from the six largest assemblages.
The final index reported in Table 18.5 relates the weight of corrugated pottery to the number of identified bones. This is calculated for each site by dividing the total weight of corrugated pottery by the total number of identifiable bones, thus creating a value expressed as grams of pottery per identified bone. As with most of these indices, this method requires some assumptions. First, we assume that corrugated pottery was produced and broken at a constant rate (relative to population size). As the pottery is common and apparently utilitarian, this seems to be a reasonable assumption (Varien and Potter 1997; Varien and Mills 1997). Second, we assume that the rate of bone discard was constant, because the index will only operate as an indicator of bone destruction if bone accumulation was originally similar from site to site. This assumption may be invalid, because, as we will show, there is significant variation in the percentage of species from site to site, and it is therefore possible that the pottery:bone index reflects both hunting intensity as well as degree of bone destruction.
Analysis of the indices reported in Table 18.5 does not reveal any clear trends. Mad Dog Tower appears to have the poorest preservation (highest percentage of unidentifiable bone; lowest average fragment length; highest percentage of burnt bone; greatest pottery to bone ratio), and this, coupled with small sample size, makes the assemblage of little value for further study. There are no good correlations of indices for other sites. For example, Lillian's Site has a low average fragment length and a higher-than-average percentage of burnt bone, but the distal:proximal ratio suggests better preservation than at other sites, and the pottery to bone ratio is in the middle of the range. Similarly, Saddlehorn Hamlet has a high average fragment length, a low percentage of burnt bone, and a low pottery to bone ratio, but a relatively high percentage of unidentified bone and a strongly imbalanced distal to proximal ratio. Data such as these suggest that the indices are probably not sensitive enough to reveal relatively minor differences in preservation between most sites, with the exception of Mad Dog Tower, which is exceptionally poorly preserved. We will therefore assume that preservation has not differentially affected the assemblages of the other 12 sites.
Intrasite Variation
Although sampling methods and excavation methods were consistent from site to site, it is worth asking whether there is significant intrasite variation in assemblages, because this could potentially affect the total assemblage recovered from the site. Intrasite variation might result from cultural practices (for example, different disposal methods for different species) or from preservation conditions (for example, different conditions inside and outside structures). It is also possible that different types of deposits (for example, midden vs. kiva floor) would contain different faunal assemblages. The relatively small sample sizes make intrasite analysis difficult. As a preliminary step in this analysis, assemblages from Castle Rock Pueblo, Kenzie Dawn Hamlet, and Stanton's Site are examined. Using provenience information supplied by the excavators, it was possible to assign most bones to a structure or to an area outside the structures. For each of these areas we calculated the number of bones in three categories--turkey plus large bird, cottontail (the most common small mammal), and deer plus medium artiodactyl. The data are presented in Table 18.6.
Table 18.6 demonstrates that the overall pattern of relative frequency of the major taxa is maintained in most types of contexts. For example, the site assemblage for Castle Rock Pueblo contains roughly equal amounts of cottontail and turkey plus large bird, and this pattern is found in most areas of the site. Similarly, the site assemblage for Stanton's Site is dominated by turkey and large bird, and this relationship holds for every subarea within the site. One consistent difference between assemblages from inside structures and those from outside structures is that turkey and large bird occurs in higher percentages in the exterior deposits than in the interior deposits. Regardless of the overall percentage of turkey and large bird in the site as a whole, it is always relatively more frequent in exterior deposits. We do not have a good explanation for this phenomenon at present, although it is possible that turkey is associated with permanent domestic occupation and the bones are often deposited outside structures, whereas some cottontail bones might be deposited inside structures after permanent residence has ended but while the structure is still being visited casually during horticultural or hunting activities (Mark Varien, personal communication 1993). Larger samples from well-defined depositional contexts will be required to explore this further. However, it seems unlikely that significant variation between assemblages was caused by sampling different types of deposits.
Intersite Variation
Variation in the composition of faunal assemblages results from a combination of causes and factors. Very generally, these may include the availability of fauna around a site; decision making by the hunters and trappers about what to hunt and when and where to do so; methods of processing and bone disposal; culture change; preservation conditions; the addition of fauna by noncultural means; and excavation and recording methods. Virtually all of these factors might play a role in causing variation between sites in the Sand Canyon locality. However, excavation and recording methods were consistent between sites, and, as suggested above, preservation is also unlikely to have had a differential impact on the various sites. The first step in this analysis is therefore to consider the differences between sites in the study group, including the time at which the site was occupied, the location of the site in relation to environmental features, and the location of the site in relation to other settlements (Table 18.7).
For purposes of the following discussion, sites are divided into groups based on location in lower or upper Sand Canyon. Lower canyon sites are found either toward the southern end of Sand Canyon (Mad Dog Tower and Saddlehorn Hamlet) or in McElmo Canyon close to the entrance to Sand Canyon (Castle Rock Pueblo). Elevations of these sites are about 300 m lower than elevations of most of the other sites discussed in this paper. The sites date to the thirteenth century, and the two largest bone assemblages in the lower canyon (Saddlehorn Hamlet and Castle Rock Pueblo) date to the second half of that century.
The upper canyon area includes the mesa-top sites, cliff/talus-slope/bench sites, and Sand Canyon Pueblo. The mesa-top sites (Shorlene's Site, Roy's Ruin, Lillian's Site, G and G Hamlet, Troy's Tower, and Kenzie Dawn Hamlet) occur above the canyon rim at the north end of Sand Canyon. Most of these sites were occupied into the thirteenth century but were abandoned as permanent residences by A.D. 1250. Some of them have one or more earlier occupations.
Cliff/talus-slope/bench sites (Catherine's Site, Lester's Site, Lookout House, and Stanton's Site) are located in the north end of upper Sand Canyon below the canyon rim. Lester's Site is located immediately adjacent to Sand Canyon Pueblo, but lies outside the boundary wall, and is therefore considered a separate site. All four sites date to the latter half of the thirteenth century.
Sand Canyon Pueblo, located at the head of Sand Canyon, was not included in the Site Testing Program, but extensive faunal collections have been made in recent excavations (Bradley 1992b) and will be used for comparative purposes. It appears that the pueblo was planned, built, and abandoned in a few decades within the second half of the thirteenth century (Bradley 1993). The mesa-top sites appear to be slightly earlier than the other sites, whereas the lower canyon sites, the cliff/talus-slope/bench sites, and Sand Canyon Pueblo are contemporaneous.
There are certain obvious consistencies in assemblages from all these sites in spite of variation in location and age. The numbers of artiodactyls, carnivores, and birds (other than turkey) are small (see Table 18.1). Cottontails account for 72 percent to 100 percent of the lagomorph assemblage, although the extreme values are from quite small assemblages and the median value is 88 percent. The rodent assemblages are usually dominated by sciurids and packrats.
Quite a lot of variation between sites appears to be the result of different percentages of rodents. As noted above, there is evidence that some of the rodents are intrusive. Excavations have demonstrated extensive burrowing, and rodents live on and in the sites today. Consequently, the variation might be caused as much by the attractiveness of a site for postabandonment rodent activity as it was by past cultural behavior. Grouping sites either by location or by time period produces no consistent patterning in the rodent assemblages.
If one excludes rodents from the analysis, variation is seen mainly as differences in the relative frequencies of turkey (including large bird) and lagomorphs. The percentage of total lagomorphs (lagomorph, cottontail, and jackrabbit together) in the total lagomorph-turkey-large bird assemblage is presented in Table 18.8 for all sites except Mad Dog Tower, with the sites organized by location.
Sand Canyon Pueblo fauna is also included in Table 18.8. Analysis of Sand Canyon Pueblo fauna has been undertaken by Walker (1990a) and by Brand (1991). Walker's results are not strictly comparable with the analyses done in this study, because identification criteria were somewhat different. In all sites reported in this study, bones were considered "identifiable" only if the element could be specified. Thus, any bone identified in this study as "large bird" can be attributed to a particular element (for example, humerus, scapula, etc.). Walker identified some fragments to class levels even if the element could not be specified. Thus, Walker's "large bird, probably turkey" category will include specimens (such as "long bone shaft fragments") which Brand would not have included because the element could not be identified. Because Brand's analysis used the same identification criteria as all other faunal analyses reported here, Brand's data were used as the comparative assemblage.
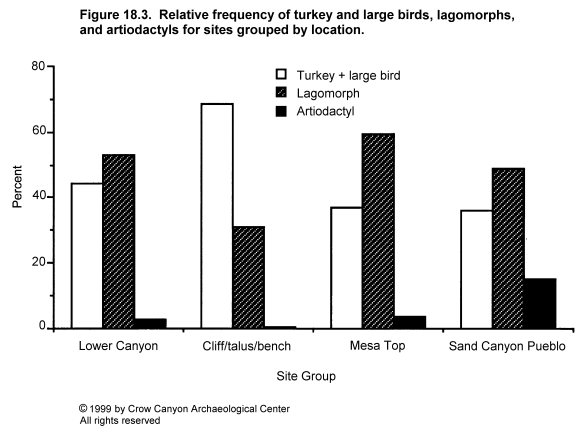
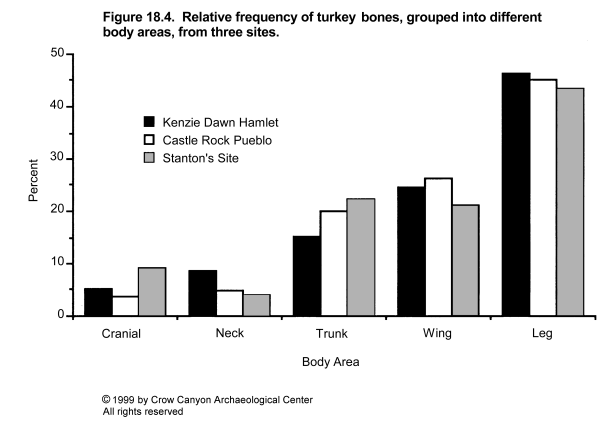
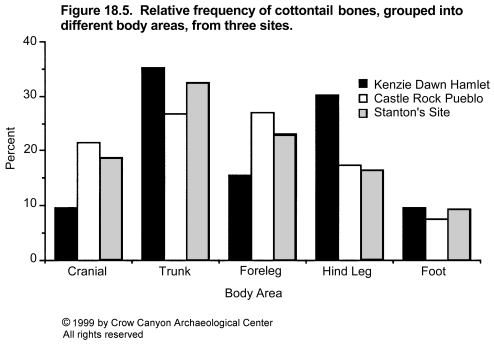
Variation in percentages of lagomorphs and turkey plus large bird is illustrated by site in Figure 18.2 and by site group in Figure 18.3, and is clearly patterned by time and location. The early-thirteenth-century settlements of the mesa top have a lagomorph to turkey plus large bird ratio of between 1:1 and 2:1. This pattern was maintained when the inhabitants of these sites abandoned their mesa-top settlements and moved to Sand Canyon Pueblo in the mid-thirteenth century. The same pattern is seen in Saddlehorn Hamlet and Castle Rock Pueblo, which are located at the lower end of Sand Canyon and in McElmo Canyon and which are contemporaneous with Sand Canyon Pueblo. However, the four cliff/talus-slope/bench sites which lie below the rim of upper Sand Canyon contain about twice as many turkey or large bird bones as lagomorph bones. In other words, the normal pattern in the Sand Canyon locality was for the number of lagomorph bones to equal or exceed the number of turkey and large bird bones by ratios of up to 2:1, but for one subset of sites the pattern is reversed. These sites are contemporaneous with Sand Canyon Pueblo and lie close to it. The implications of this will be discussed later, but it is worth noting now that these quantitative differences do not seem to be caused by variation in the overall representation of different areas of the skeleton in lagomorphs and turkeys. To illustrate this, cottontail and turkey bones from the three largest sites have been grouped into various body areas. The assemblages include sites from the mesa top (Kenzie Dawn Hamlet), the lower canyon (Castle Rock Pueblo), and the cliff/talus-slope/bench (Stanton's Site) groups. As Figure 18.4 and Figure 18.5 demonstrate, there is little variation in body part representation between the three sites.
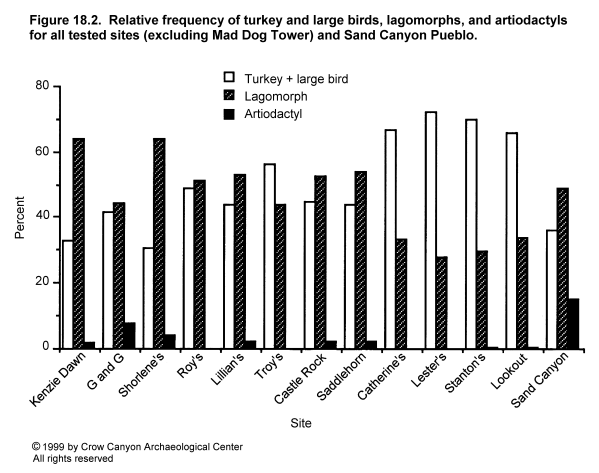{kind=link}
{kind=link}
{kind=link}
{kind=link}
Variation in the quantity of artiodactyls (principally deer) is more difficult to detect because numbers of artiodactyls are lower. Nevertheless, data in Table 18.8 and Figure 18.2 and Figure 18.3 show some regularities. The four cliff/talus-slope/bench sites consistently contain less artiodactyl bone than do other sites. Stanton's Site, which has one of the largest assemblages studied in this project, contains less than one percent artiodactyl bone, showing that this is not simply an effect of small sample size for some assemblages. Deer are more frequent in the mesa-top sites (3.1 percent of the total mesa-top assemblage) and in the lower canyon sites (averaging 2.6 percent). Sand Canyon Pueblo contains significantly more artiodactyl bone than any other site except G and G Hamlet, where the presence of a few artiodactyl bones in a very small assemblage creates a fairly high percentage. Although these differences may appear slight in terms of percentages of the number of identified bones, it is worth remembering that a small increase in the number of deer killed would have a substantial effect on the quantity of meat available, because a deer weighs 50 to 100 times as much as a cottontail.
Thus, there appears to be a fairly well defined pattern of faunal variation which relates to site location, time of occupation, and settlement size. Prior to the founding of Sand Canyon Pueblo, mesa-top sites were characterized by relatively high lagomorph to turkey ratios and a fairly small percentage of artiodactyls. There is no information on sites in other locales. This pattern is continued in the lower canyon sites dating to the later thirteenth century, and this suggests that the common Pueblo III pattern in this area was an emphasis on small mammal procurement, raising domestic turkey, and infrequent artiodactyl hunting. This is the same pattern that one sees in earlier Puebloan sites in this region (e.g., Neusius 1986) but with an increased emphasis on raising turkey.
The growth of Sand Canyon Pueblo seems to coincide with the abandonment of many mesa-top sites as habitations and the development of the smaller cliff/talus-slope/bench sites within upper Sand Canyon. Sand Canyon Pueblo fauna is reasonably similar to the mesa-top sites and to the lower canyon sites, in that the lagomorph to turkey ratio is high. The main difference is that artiodactyls are more important at Sand Canyon Pueblo than in other locations.
Explanation of Intersite Variation
For the tested sites, there are three interrelated questions which require further discussion: (1) Why do the cliff/talus-slope/bench sites of the late thirteenth century differ from other sites on the mesa top and the lower canyon? (2) Why do all late-thirteenth-century sites contain much less deer bone than the contemporaneous Sand Canyon Pueblo? (3) What was the relationship between the cliff/talus-slope/bench sites and Sand Canyon Pueblo? The analysis of the tested sites also raises other questions in relation to earlier periods. All of the tested sites have much higher percentages of turkey bone than Pueblo I and Pueblo II sites in the same general region (e.g., Neusius 1986; Walker 1989). The intensification of turkey production has been addressed in a separate study (Munro 1994).
When dealing with species which were of dietary importance, investigation of variation should commence with a consideration of environmental or economic factors. Speth and Scott (1989) have commented on the increased importance of large mammals in aggregated communities in the Southwest. They propose that small mammals may be "hunted out" around large settlements, and that as hunting trips become longer, it makes sense to switch to larger prey. In addition, larger settlements may be able to organize communal hunts more efficiently. This seems a plausible hypothesis for the increase of artiodactyls at Sand Canyon Pueblo, although one might have expected a similar increase in deer at Castle Rock Pueblo, which is one of the largest late-thirteenth-century sites in McElmo canyon. However, Castle Rock Pueblo was not nearly as large as Sand Canyon Pueblo, and overexploitation of local game may not have occurred. If long-range hunting was being practiced at Sand Canyon Pueblo, we would expect to see evidence for this in the species being hunted and in the parts of the animal being brought back to the site. Both Walker (1990a) and Brand (1991) document occasional use of bighorn sheep. Walker's analysis indicates that bighorn bones were often modified into artifacts, which might suggest trade rather than long-range hunting as a method of acquiring bighorn products. Bighorn bones identified by Brand are primarily from the head and feet, which are not prime meat-bearing bones and might be expected to be abandoned at a kill, especially if the meat was to be transported long distances. On the other hand, bones such as ribs and vertebrae, which might be transported, are unlikely to be identified to species. Brand (1991) also identified a few pronghorn antelope bones, and Walker (1990a) speculated that a large artiodactyl rib might be from bison. These would also indicate long-distance acquisition of meat.
Counts of elements identified as deer and medium artiodactyl by Brand (1991) do not demonstrate any particular selectivity in elements, and all areas of the body were brought back to Sand Canyon Pueblo. When compared with the total artiodactyl assemblage from all of the tested sites, bones from the trunk are slightly more common at Sand Canyon Pueblo and foot bones are slightly less common (Table 18.9). Although this may suggest more frequent discard of foot bones before transportation of deer to Sand Canyon Pueblo, the differences are relatively minor, and it is difficult to support the long-range hunting hypothesis.
Continuing with environmental/economic hypotheses, the reduction in lagomorphs and increase in turkey at the cliff/talus-slope/bench sites might be explicable in terms of the location of these sites within the canyon, but this does not seem very likely. All of these sites are very close to the mesa top, and their catchment zones would overlap to a large extent with the areas around the slightly earlier mesa-top sites in which lagomorphs were common. Furthermore, Sand Canyon Pueblo is located very close to the cliff/talus-slope/bench sites, and at this site, lagomorphs continued to outnumber turkey. Thus, site location in relation to the surrounding natural environment does not seem to account for the cliff/talus-slope/bench assemblages.
Hypotheses based on economic change are not strongly supported in the above analyses. Hypotheses derived from models of social organization will therefore be considered next. First, it can be suggested that inhabitants of various sites were excluded from access to certain resources by the inhabitants of other sites, notably those of Sand Canyon Pueblo. In this scenario, the inhabitants of the cliff/talus-slope/bench sites would have been excluded almost completely from hunting grounds outside of the canyon. Lagomorphs would have been obtained in relatively small numbers from within the canyon, and access to deer would have been minimal. As a result, they were forced to raise increased quantities of turkey to compensate for a lack of hunted meat. The inhabitants of the lower canyon sites were far enough away from Sand Canyon Pueblo (here interpreted as politically dominant) to continue the hunting patterns of the early thirteenth century (as exemplified by the mesa-top sites). However, no lower canyon sites, including Castle Rock Pueblo, were sufficiently large enough to exclude others from deer-hunting territory, and there was no rise in the percentage of deer. This hypothesis would see the inhabitants of the smaller sites near Sand Canyon Pueblo as socially/politically disadvantaged. At Sand Canyon Pueblo itself, the ratio of lagomorphs to turkey remained similar to the earlier pattern, but control of deer-hunting territory allowed greater numbers of deer to be killed. Essentially, this hypothesis proposes that aggregated communities were able to intimidate their neighbors and gain monopolies over certain resources.
Alternatively, the inhabitants of the cliff/talus-slope/bench sites may have been fully integrated into the social life of Sand Canyon Pueblo. This hypothesis considers individual sites to have been part of larger communities (e.g., Rohn 1989), and would see one large community at the head of Sand Canyon in the late thirteenth century, with a population concentration in Sand Canyon Pueblo and outlying households represented by cliff/talus-slope/bench sites. In this hypothesis, the inhabitants of the small outlying sites would be integrated into the large community. (The lower Sand Canyon community at this time would have centered around Castle Rock Pueblo, but social organization was sufficiently similar to earlier Pueblo III settlements that no change in the animal economy occurred.)
Hunting was of considerable social and ritual importance in ethnographically known Southwestern societies (Gnabasik 1981), and it seems unlikely that Puebloans in prehistoric times would have reduced their participation in this important activity voluntarily. Hunting is often linked to males, but communal rabbit hunts in the Southwest sometimes included both men and women. The ethnographic literature concentrates on the large communal hunts, and there is less information about snaring and trapping individual cottontails and jackrabbits. On the other hand, deer (and other artiodactyl) hunting appears to have been almost exclusively a male activity. For turkeys, the literature again tends to concentrate on the importance of turkey feathers in ritual paraphernalia and does not discuss stock-raising or the position of the turkey in the domestic economy. However, the act of procuring turkey meat does not seem to have been accorded any special significance.
One could argue that Sand Canyon Pueblo assemblages are to a greater extent the product of hunting undertaken for ritual and social purposes, rather than for purely economic needs. Deer hunting was of particular significance in most Southwestern societies, and was often attended by numerous ritual events. The relatively high cottontail:turkey ratio (when compared with the nearby contemporaneous cliff/talus-slope/bench sites) might suggest greater emphasis on communal rabbit hunting, again a public hunt with important ritual and social functions. Sand Canyon Pueblo is distinguished by the importance and prominence of its public architecture (Bradley 1993), and this interpretation of the hunting procedures reflects the architectural reconstruction. Assuming that greater ritual emphasis has an integrative function in larger communities (Hegmon 1989; Johnson 1989), the higher percentage of deer at Sand Canyon Pueblo might reflect its role in a more highly organized religious system. If so, one would expect to see more deer associated with ritual structures at Sand Canyon Pueblo.
There is some evidence from Sand Canyon Pueblo that architectural blocks with high kiva:room ratios have higher amounts of medium artiodactyls (mainly deer) and lagomorphs (mainly cottontail) than blocks with lower kiva:room ratios. Table 18.10 uses data from Walker (1990a:Table 8) to compare percentages of major taxa in different architectural blocks. Kiva: room ratios are taken from Bradley (1992b). As noted before, Walker's methods of identification were different from those used in the other faunal analyses discussed in this paper, so the faunal data presented in Table 18.10 cannot be compared directly with data generated by our analyses. However, the trends in Walker's data clearly demonstrate that the highest percentage of turkey plus large bird and the lowest percentage of deer occur in the architectural block with the fewest kivas, and the highest percentages of deer occur in architectural blocks with the most kivas. This suggests that kivas at Sand Canyon Pueblo may have been focal points for feasting on hunted animals. These topics will be explored in more detail when a full analysis of Sand Canyon Pueblo fauna is complete.
One might also argue that the cliff/talus-slope/bench sites display a domestic pattern rather than a public pattern. Turkeys were presumably raised in and around households by families, and one suspects that this may have been undertaken more by women than by men. The very low percentages of deer at cliff/talus-slope/bench sites suggest minimal participation in public hunts. The relatively low cottontail:turkey ratios could be the result of local trapping of cottontails, carried out by individuals rather than by larger social groups. If, as suggested elsewhere (Varien et al. 1992; Lipe 1992b), the inhabitants of the mesa-top sites moved from the mesa top to Sand Canyon Pueblo and retained ownership of the fields on the mesa top, they may have continued to organize rabbit hunts and deer hunts on the mesa top.
Another hypothesis is that there were specialized economic relations between Sand Canyon Pueblo and the cliff/talus-slope/bench sites, with the latter specializing in the production of turkey meat and feathers for Sand Canyon Pueblo, and receiving other products in exchange. However, given the probable importance of deer hunting as an activity, it seems unlikely that the inhabitants of the cliff/talus-slope/bench sites would have given up that activity, even if they could trade for deer meat with their neighbors. Unless further evidence for exchange appears from the artifact assemblages, this hypothesis can probably be discarded.
The first and second hypotheses present two views of the relationship between Sand Canyon Pueblo and the nearby sites. The first hypothesis views the inhabitants of the cliff/talus-slope/bench sites as being excluded from important economic resources by a powerful neighbor. The second hypothesis sees them as integrated into certain community-wide ritual and ceremonial events which were based in Sand Canyon Pueblo, and which may have included both communal hunts and feasts. Both of these hypotheses presume the presence of people in Sand Canyon Pueblo who were able to organize the procurement and distribution of more deer meat than members of other communities, including the lower canyon, late-thirteenth-century group and the slightly earlier, mesa-top community. Unfortunately, the faunal remains from Sand Canyon Pueblo have not yet been analyzed in sufficient detail to allow more discussion of possible modes of social organization at that site and in the surrounding community of cliff/talus-slope/bench sites.
Conclusions
The faunas from the 13 tested sites show numerous similarities, as one would expect when comparing sites separated by no more than 10 km in space and less than two centuries in time. The main components of the assemblages are lagomorphs (mainly cottontail rabbits), domestic turkey, and artiodactyls (mainly deer). Amphibians and reptiles are assumed to be intrusive in most cases. Analysis of rodents suggests that a greater proportion of their remains are intrusive than are the remains of cottontail, but the overall importance of rodents cannot be determined. It seems likely that they were trapped or hunted on some occasions, but no consistent patterns of intersite variation were observed.
Preservation conditions vary somewhat between sites, although only one site (Mad Dog Tower) has significantly worse preservation. At all sites, bone has been damaged by a variety of natural and cultural factors, including carnivore damage (rare), rodent gnawing, weathering, trampling, and burning. The degree of damage seems similar from site to site, and therefore the assemblages should be broadly comparable. There is relatively little intrasite variation.
Variation is seen most easily when the relative values of the major taxa are considered. The turkey:lagomorph ratio is fairly constant, except at cliff/talus-slope/bench sites within the upper part of Sand Canyon, where turkey is twice as common as lagomorphs. These sites are late in the sequence and are at least partly contemporaneous with Sand Canyon Pueblo, located at the head of Sand Canyon. These sites are also distinguished by very low percentages of artiodactyl bones. Sites located in lower Sand Canyon, McElmo Canyon, and on the mesa top around upper Sand Canyon have turkey:lagomorph ratios similar to those at Sand Canyon Pueblo, and they have artiodactyl percentages that are intermediate between those documented for Sand Canyon Pueblo and the upper Sand Canyon cliff/talus-slope/bench sites. The distinctive nature of the latter is explained tentatively by the dominant social and political position of Sand Canyon Pueblo, but this explanation requires further evidence from faunal and other analyses.
Models of social organization in aggregated communities in the Southwest range from hierarchical elites to egalitarian lineages. The primary reason that analysis of social organization has been so controversial is the lack of middle-range models which link material evidence to past social conditions (e.g., Lightfoot and Upham 1989). Zooarchaeology provides the methods for examining regular acquisition of a range of desirable foods, and social organization can potentially be examined through such information (e.g., Crabtree 1990). Future work on intrasite variation at more extensively excavated sites will attempt to utilize such methods.
Acknowledgments
We thank the Crow Canyon Archaeological Center for inviting us to participate in this research and for providing funding for some of the analyses. We especially thank Karen Adams, Bruce Bradley, Bill Lipe, and Mark Varien for time, information, and advice. Funding was provided to Driver by the Social Sciences and Humanities Research Council of Canada. The Department of Archaeology, Simon Fraser University, provided laboratory facilities for the analyses reported here.